P5 – Dissecting the trafficking events leading to herpesvirus secondary envelopment
Principal Investigators
Prof. Dr. rer. nat. Kay Grünewald
Center for Structural Systems Biology, Hamburg
Structural Cell Biology of Viruses
Prof. Dr. rer. nat. Jens Bosse
Center for Structural Systems Biology, Hamburg
Quantitative Virology
Project Summary
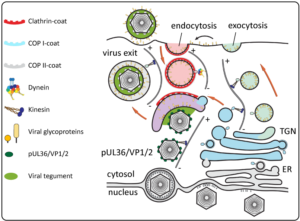
The nine human herpesviruses present medically highly relevant, double-stranded and enveloped DNA viruses. Due to their long co-evolution with their hosts, herpesviruses are also ideal ‘tools’ to study cell biological events they hijack along their ‘life’-cycle. Virus entry involves fusion of the virus envelope with the plasma membrane or endosomal membranes mediated by a multi-component fusion machinery of several viral glycoproteins. Cytosolic capsids are then transported to nuclear pores, where the viral genome is injected into the nucleoplasm. Thereafter, viral genomes replicate and are packaged into newly assembled capsids1. Progeny capsids egress the nucleus by budding at the inner nuclear envelope (primary envelopment) and subsequent fusion of the formed perinuclear vesicles with the outer nuclear envelope2. In a process termed secondary envelopment, cytosolic capsids bud into cellular vesicles carrying viral glycoproteins (see Fig.). The process is mediated by a network of protein interactions of viral proteins (tegument). This results in enveloped viral particles inside transport vesicles that finally fuse with the plasma membrane to release virions. Secondary envelopment is an essential step in virion assembly, and hence presents an ideal target for antiviral interference. However, it is currently unclear which cellular membranes are used by the virus for envelopment. Early reports suggested that envelopment happens at trans-Golgi-derived membranes (COP1-coated vesicles) while more recent studies argue that early endosomes (clathrin-coated vesicles) might be the source. These studies are often based on few specific markers that serve as surrogates for the vesicle origin. However, it has been shown that herpesviruses induce dramatic changes to the cellular secretory machinery and these changes likely also change the distribution and abundance of cellular proteins typically used as cellular compartment markers. It emerges that, secondary envelopment might happen at the crossroad of several vesicle pathways3. More generally, it has become clear that lipids determine the fate of membranes and marker proteins often only temporally associate with them which is a problem for determining vesicle provenance. Binary labelling of vesicle populations based on a limited number of markers likely does not reflect the nature of the cellular secretory system which is of more gradual nature with myriads of mixing and de-mixing events. Own preliminary data suggest that individual viral glycoproteins take different pathways through the secretory system involving dynein and kinesin motor complexes for microtubule-based transport before coming together and mediating secondary envelopment of herpesvirus capsids (see Fig.).
Here, we aim to characterise the provenance of membranes and the pathway of viral glycoproteins involved in secondary envelopment of Herpes simplex virus 1 (HSV-1), the Alphaherpesvirinae prototypic species. HSV-1 carries 12 different glycoproteins on its surface. Employing a functional approach combining single particle tracking and native electron cryo tomography (cryo-ET) plus statistical data on the localisation and behaviour of vesicle populations we aim to reveal their origin without requiring cellular markers to deduce the membrane identity.
References
1. (2015) Remodeling nuclear architecture allows efficient transport of herpesvirus capsids by diffusion. Proc Natl Acad Sci U S A 112:E5725-33. Abstract
2. (2015) Structural Basis of Vesicle Formation at the Inner Nuclear Membrane. Cell 163:1692-701. Abstract
biricu I, Maurer UE, Grünewald K (2013) Characterization of herpes simplex virus type 1 L-particle assembly and egress in hippocampal neurones by electron cryo-tomography. Cell Microbiol 15:285-91. Abstract
4. (2020) The prefusion structure of Herpes simplex virus glycoprotein B. Sci Adv 6: eabc1726. Abstract
. Abstract